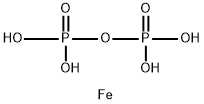
As a common organic iron supplement, ferrous gluconate exhibits superior properties compared to inorganic iron (e.g., ferrous sulfate), including better water solubility, lower intestinal irritation, and higher bioavailability. Its regulatory effect on the intestinal microbiota is not achieved through direct "selective killing" or "targeted proliferation" but rather through the controllable release of iron ions, mild influence on the intestinal microenvironment, and compatibility with the iron metabolism needs of microorganisms—indirectly improving the balance of microbiota structure and function. This can be analyzed in detail from three aspects: mechanism of action, impact on microbiota composition, and associated physiological effects.
I. Mechanism of Action: Microenvironment Regulation Centered on "Gentle Iron Supplementation"
The survival and metabolism of the intestinal microbiota are highly dependent on iron—an essential cofactor for microbial respiratory chain enzymes (e.g., cytochromes), nucleic acid synthases (e.g., ribonucleotide reductases), and antioxidant enzymes (e.g., catalases). Different microbial groups vary in their iron acquisition capabilities and demand thresholds. The regulatory effect of ferrous gluconate on the microbiota originates first from its unique iron release pattern:
On one hand, ferrous gluconate dissociates slowly into gluconate ions and divalent iron ions (Fe²⁺) in the intestine (especially in the ileum and colon, where the pH is neutral), avoiding the problem of sudden local iron concentration spikes caused by the rapid release of inorganic iron. Excess free iron disrupts the intestinal redox balance, inducing overproduction of reactive oxygen species (ROS), which in turn damages beneficial bacteria sensitive to oxidative stress (e.g., Bifidobacterium, lactic acid bacteria). The slow-release property of ferrous gluconate maintains intestinal iron concentration within an optimal range—"meeting the basic iron needs of microorganisms without causing oxidative damage"—creating a stable iron-nutrient microenvironment for the microbiota.
On the other hand, the organic ligand (gluconate) of ferrous gluconate forms a weak coordination bond with Fe²⁺, reducing the binding of Fe²⁺ to antinutritional factors in the intestine (e.g., tannic acid, phytic acid) and improving iron bioavailability. This not only provides the host with an adequate iron source but also enables intestinal microorganisms to acquire iron more efficiently, preventing microbiota metabolic disorders caused by "iron deficiency." For example, when the host is iron-deficient, competition for iron in the intestine intensifies. Some pathogenic microorganisms (e.g., Escherichia coli, Salmonella) secrete siderophores (e.g., enterobactin) to seize iron sources, squeezing the living space of beneficial bacteria. Supplementation with ferrous gluconate alleviates this "iron competition pressure" and creates conditions for beneficial bacteria to utilize iron.
Additionally, ferrous gluconate indirectly affects the microbiota by regulating intestinal pH. Intestinal pH is a key factor regulating microbiota distribution (e.g., lactic acid bacteria prefer acidic environments, while Clostridium prefers neutral to slightly alkaline environments). As a weakly acidic group, gluconate slightly lowers local intestinal pH. Although this is insufficient to alter the overall intestinal acid-base balance, it provides a more suitable microenvironment for acid-producing beneficial bacteria (e.g., lactic acid bacteria), indirectly promoting their proliferation while inhibiting the growth of some harmful bacteria sensitive to acid (e.g., Clostridium perfringens).
II. Impact on Intestinal Microbiota Composition: Optimizing Microbiota Structure and Diversity
Under optimal iron supply and microenvironment regulation, ferrous gluconate optimizes the intestinal microbiota structure by "supporting beneficial bacteria, inhibiting harmful bacteria, and maintaining microbiota diversity":
For beneficial bacteria, supplementation with ferrous gluconate significantly promotes the proliferation of core beneficial bacteria such as Bifidobacterium and lactic acid bacteria. These microbial groups have relatively "gentle" iron metabolism systems and rely on stable intestinal iron concentrations for metabolic activities. For example, Bifidobacterium requires iron to synthesize lactase (which breaks down lactose to produce lactic acid), while lactic acid bacteria need iron to maintain cell wall integrity and acid-producing capacity. Studies have shown that after 4–8 weeks of ferrous gluconate supplementation in iron-deficient individuals, the number of Bifidobacterium in feces increases by 2–3 orders of magnitude, and the proportion of lactic acid bacteria also rises significantly—directly attributed to the continuous iron source and weakly acidic microenvironment provided by ferrous gluconate.
For harmful bacteria, ferrous gluconate acts mainly through "competitive inhibition" and "oxidative stress regulation." On one hand, the slowly released Fe²⁺ is preferentially utilized by beneficial bacteria, reducing the amount of iron available to harmful bacteria (e.g., Salmonella, Shigella) and impairing their virulence. For instance, the invasiveness of Salmonella depends on iron-mediated virulence gene expression; when iron supply is insufficient, its ability to adhere to intestinal epithelial cells decreases significantly. On the other hand, ferrous gluconate prevents ROS bursts caused by excess free iron, reducing the overproliferation of harmful bacteria triggered by oxidative stress (some harmful bacteria are more tolerant to ROS than beneficial bacteria, and excess ROS may instead promote their growth).
Notably, the role of ferrous gluconate in maintaining microbiota diversity is particularly critical. In iron-deficient states, intestinal microbiota diversity often decreases due to "iron-dependent microbiota dominance" (e.g., increased proportion of iron-tolerant harmful bacteria). Supplementation with ferrous gluconate restores microbiota species richness by balancing iron supply, avoiding microbiota imbalance caused by the overproliferation of a single microbial group. This is a key feature distinguishing it from some antibiotics or high-potency prebiotics, which may cause microbiota structural simplification due to their strong effects.
III. Associated Physiological Effects: From Microbiota Regulation to Intestinal Health Improvement
By regulating the intestinal microbiota, ferrous gluconate further improves intestinal barrier function, metabolic function, and immune function, forming a positive cycle of "iron supplementation → microbiota regulation → intestinal protection":
In terms of intestinal barrier function, proliferated Bifidobacterium and lactic acid bacteria secrete short-chain fatty acids (SCFAs, e.g., acetic acid, propionic acid, butyric acid), which promote the expression of tight junction proteins (e.g., occludin, claudin) in intestinal epithelial cells and strengthen the intestinal physical barrier. Meanwhile, metabolites of beneficial bacteria inhibit the entry of endotoxins (e.g., lipopolysaccharide, LPS) secreted by harmful bacteria into the bloodstream, reducing intestinal inflammatory responses. Studies have shown that iron-deficient anemic patients supplemented with ferrous gluconate exhibit significantly increased fecal SCFA levels and decreased serum endotoxin levels—directly linked to optimized microbiota structure.
In terms of metabolic function, the intestinal microbiota is an important "partner" in host metabolism. The microbiota regulated by ferrous gluconate participates more efficiently in nutrient metabolism. For example, Bifidobacterium promotes the breakdown of dietary fiber to produce more beneficial metabolites; lactic acid bacteria assist the host in absorbing B vitamins (e.g., B12, folic acid), and these vitamins interact synergistically with iron to further improve host hematopoietic function. This "microbiota-host metabolic interaction" means that the iron-supplementing effect of ferrous gluconate extends beyond iron itself—it also enhances overall nutrient utilization efficiency through microbiota optimization.
In terms of immune function, the intestinal microbiota is a key regulator of intestinal mucosal immunity. Beneficial bacteria promoted by ferrous gluconate regulate immune factor secretion through interactions with intestinal immune cells (e.g., dendritic cells, T cells). For example, lactic acid bacteria induce dendritic cells to secrete anti-inflammatory cytokines such as IL-10, inhibiting excessive inflammation; Bifidobacterium promotes the proliferation of Treg cells, enhancing intestinal immune tolerance. For individuals with impaired immune function due to iron deficiency, ferrous gluconate indirectly enhances the immune defense capacity of the intestinal mucosa by regulating the microbiota, reducing the risk of intestinal infections.
In essence, the regulatory effect of ferrous gluconate on the intestinal microbiota lies in the precise compatibility between "the gentle properties of organic iron" and "the iron needs of the intestinal microbiota." Its slowly released iron ions not only meet microbial metabolic needs but also avoid microenvironment imbalance. By supporting beneficial bacteria, inhibiting harmful bacteria, and maintaining microbiota diversity, it indirectly improves intestinal barrier, metabolic, and immune functions. This regulatory effect differs from the "single iron supplementation" of traditional iron supplements; instead, it combines "iron supplementation" with "intestinal health maintenance," providing a more holistic nutritional support solution for iron-deficient individuals—especially those with concurrent intestinal microbiota disorders.